NMN is a popular supplement in the longevity circles. But can it boost height gain? IT’s also possible that Niacin supplementation could increase if NMN can as Niacin and NMN are related. NMN is derived from Niacin and NMN is part of the Vitamin B3 family. But NMN likely only will increase height if taken early in development but it is also possible that NMN will only increase height if one is deficient.
“Sirtuin 6 (Sirt6) is a mammalian homologue of NAD1-dependent histone deacetylase Sir2. Although Sirt6-KO mice exhibit growth retardation”
“In Sirt6-KO mice exhibited impaired proliferation and differentiation of chondrocytes, reduced expression of Indian hedgehog (Ihh), and a senescent phenotype. When Sirt6 was knocked down in chondrocytes in vitro, expression of Ihh and its downstream genes were reduced.”
“stress-response and chromatin-silencing factor Sir2, a yeast sirtuin, is a NAD1-dependent histone deacetylase and is involved in various nuclear actions”
“Sirt6 inhibits inflammation by suppressing NF-kB target molecules via interaction with the RelA subunit of NF-kB and by deacetylating H3K9 at target promoters”
“Sirt6–null mice exhibit dwarfism”
Here’s the part of the study about NMN:
“to study the effect of gain of Sirt6 activity on chondrocyte differentiation, nicotinamide mononucleotide (NMN), a key NAD1 intermediate, was administered to ATDC5 cells. NMN facilitates activity of sirtuins through enhancement of NAD biosynthesis. NMN administration enhanced Col10a1 expression together with increased expression of Ihh and Gli1, and these effects were clearly abrogated by Sirt6 silencing”
“NMN treatment enhanced Col10a1, Ihh and Gli1”
This study here:
Nicotinamide Mononucleotide Alleviates Osteoblast Senescence Induction and Promotes Bone Healing in Osteoporotic Mice
says that NMN can “NMN supplementation attenuates senescent cell induction in growth plates”<- so it’s possible NMN can keep the growth plates open for longer. Couldn’t get the full study though.
“NMN supplementation stimulated osteogenesis of endogenous MSCs, and protected bone from aging and irradiation induced damage in mice. Mechanistically, we found that NMN treatment upregulated SIRT1”
It should be noted that NMN is available in foods and laboratory animals tend not to have the most diverse diets. So we don’t know if enough NMN is gotten from foods.
There are studies in mice/rats that show that swimming increases bone length. But the kind of swimming that rats due is different than the kind of swimming that humans do.
The kind of swimming that rats/mice do is more akin to doggy paddling style swimming. This is doggy paddling that kind of mimics what the rat is doing but I think the rat style of doggy peddling is more upright.
This style may grow the arms but not the legs as when you’re upright there’s more load on the bones and more torsion. And I believe that torsion is why tennis players get longer arms and also some weightlifters. There’s more torsional load on the arms than the legs as arms can grip things. Hence why there’s more anecdotal evidence of arms growing in length rather than legs but that does not mean mechanisms to grow in the legs do not exist.
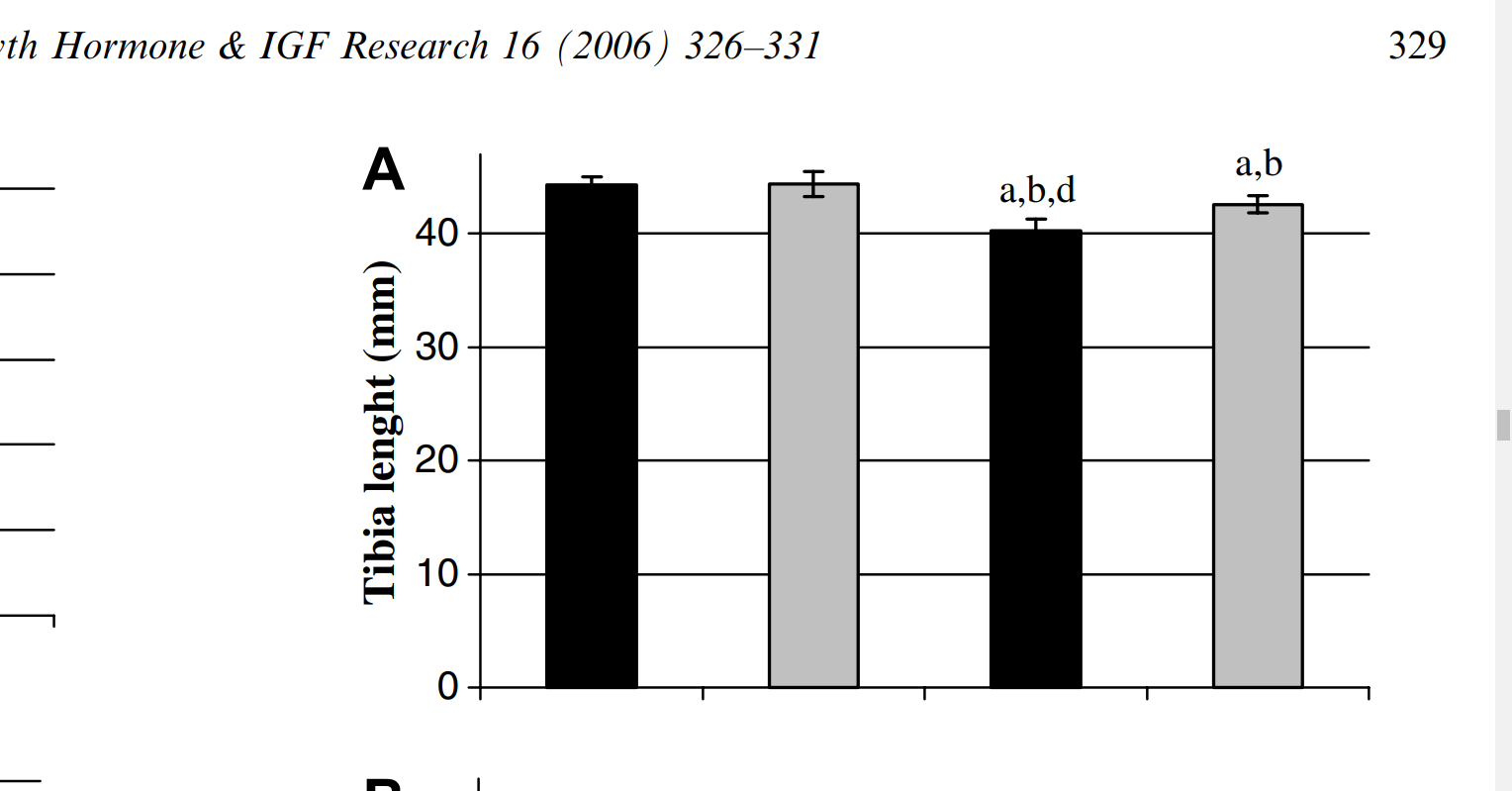
Effects of swimming training on bone mass and the GH/IGF-1 axis in diabetic rats
“The aim of this study was to examine the influence of moderate swimming training on the GH/IGF-1 growth axis and tibial mass in diabetic rats. Male Wistar rats were allocated to one of four groups: sedentary control (SC), trained control (TC), sedentary diabetic (SD) and trained diabetic (TD). Diabetes was induced with alloxan (35 mg/kg b.w.). The training program consisted of a 1 h swimming session/day with a load corresponding to 5% of the b.w., five days/week for six weeks. At the end of the training period, the rats were sacrificed and blood was collected for quantification of the serum glucose, insulin, GH, and IGF-1 concentrations. Samples of skeletal muscle were used to quantify the IGF-1 peptide content. The tibias were collected to determine their total area, length and bone mineral content. The results were analyzed by ANOVA with P < 0.05 indicating significance. Diabetes decreased the serum levels of GH and IGF-1, as well as the tibial length, total area and bone mineral content in the SD group (P < 0.05). Physical training increased the serum IGF-1 level in the TC and TD groups when compared to the sedentary groups (SC and SD), and the tibial length, total area and bone mineral content were higher in the TD group than in the SD group (P < 0.05). Exercise did not alter the level of IGF-1 in gastrocnemius muscle in nondiabetic rats, but the muscle IGF-1 content was higher in the TD group than in the SD group. These results indicate that swimming training stimulates bone mass and the GH/IGF-1 axis in diabetic rats.”
Columns: SC – sedentary control, TC – trained control, SD – sedentary diabetic, TD – trained diabetic. So the swimming groups had non-statistically significant increases in bone length between sedentary and trained control and statistically significant between sedentary diabetic and trained diabetic.
Here’s another study that finds an increase in bone length:
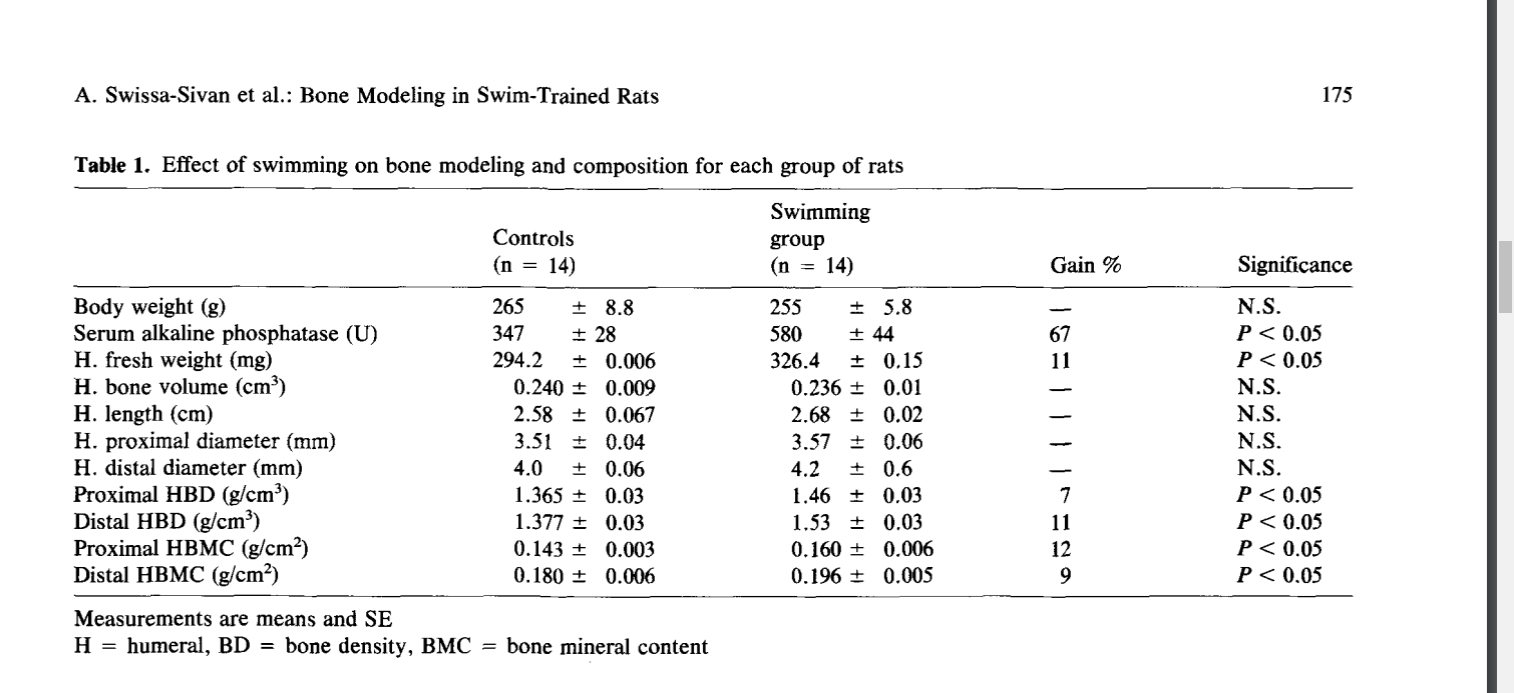
The Effect of Swimming on Bone Modeling and Composition in Young Adult Rats
“The purpose of this study was to investigate the adaptability of long bones of young adult rats to the stress of chronic aquatic exercise. Twenty-eight female Sabra rats (12 weeks old) were randomly assigned to two groups and treatments: exercise (14 rats) and sedentary control (14 rats) matched for age and weight. Exercised animals were trained to swim in a water bath (35°±1°C, 1 hour daily 5 times a week) for 12 weeks loaded with lead weights on their tails (2% of their body weight) (BW). At the end of the training period following blood sampling for alkaline phosphatase, all rats were sacrificed and the humeri and tibiae bones were removed for the following measurements: bone morphometry, bone water compartmentalization, bone density (BD), bone mineral content (BMC), and bone ions content (Ca, Pi, Mg, Zn). The results indicate that exercise did not significantly affect the animals’ body weight, bone volume, or length and diameters. However, bone hydration properties, BD, bone mass, and mineralization revealed significant differences between swim-trained rats and controls (P<0.05). Longitudinal (R1) measurement was higher by 43% for both humerus and tibia, and Transverse (R2) relaxation rates of hydrogen proton were higher by 117 and 76% for humerus and tibia, respectively; fraction of bound water was higher by 36 and 46% for humerus and tibia, respectively. BD, bone weight, and ash were higher by 13%. BMC and bone ions content were higher by 10%, and alkaline phosphatase was higher by 67%. These results indicate that long bones of young adult rats after the age of rapid growth can adapt positively to nonweight-bearing aquatic exercise. This adaptation is evident by an increase in bone mass, density, mineralization, and hydration properties.”
So perhaps upright doggy paddling is a method to induce torsional strain on relevant bones and increase height.
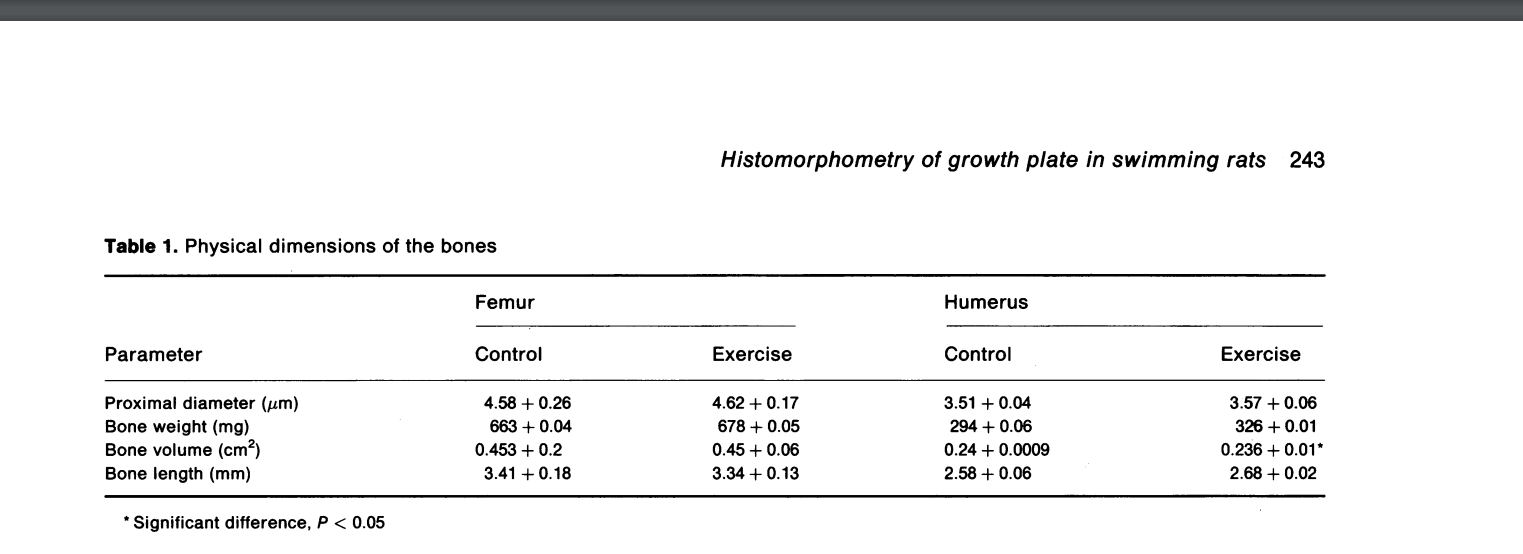
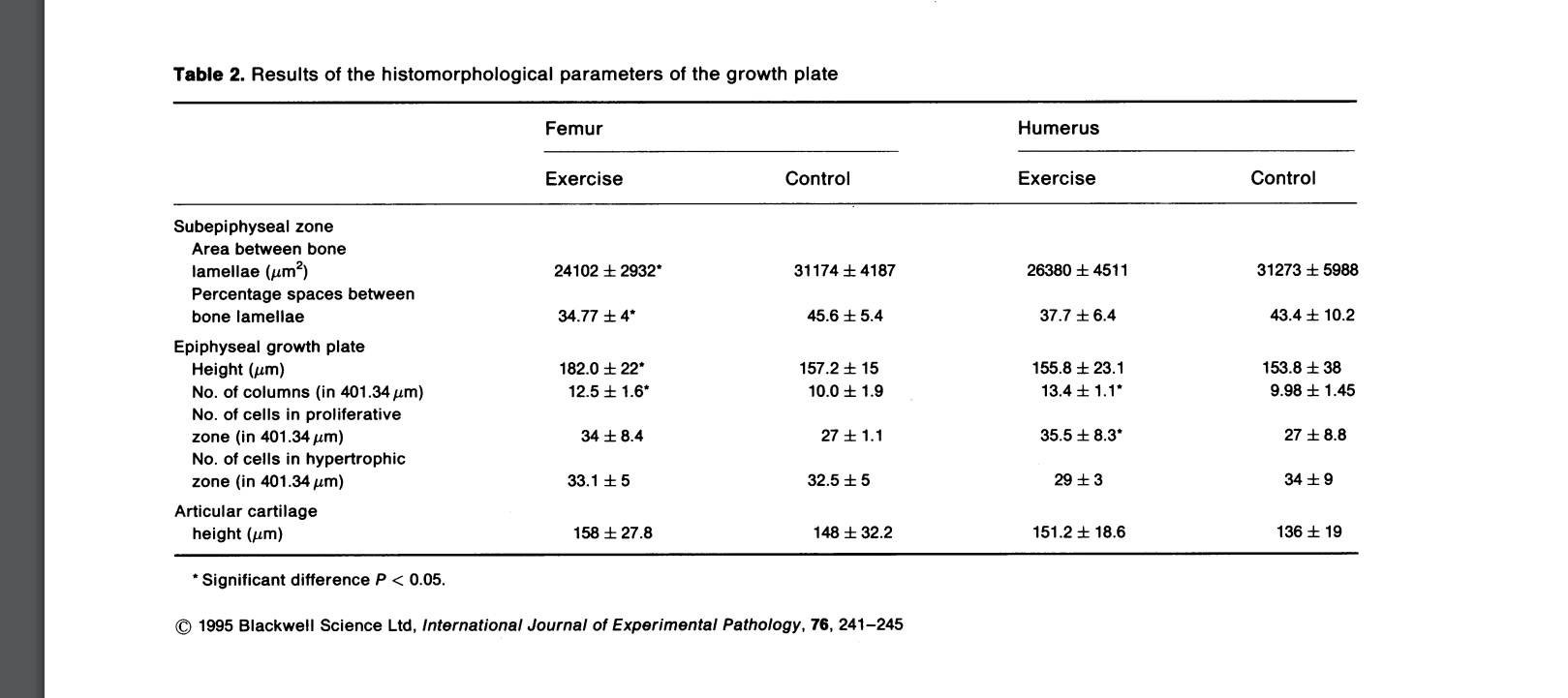
“In exercises involving running, muscle power and gravitational forces act together to affect bone mass in accordance with Wolff’s law. However, the direct effect of muscle activity on bones in non-weight-bearing activities, such as swimming, has not been explored. Previous data indicate that swimming exerts a positive effect on bone growth and development in young rats. We performed a histomorphometric study on the effect of swimming on the growth plate and subepiphyseal area of young adult rats. The experiments were carried out on 28 12-week-old albino Sabra rats. One group of 14 rats was trained to swim 1 hour/day, 5 days a week, for 12 weeks. Another group of 14 rats served as controls. The proximal femur and humerus of each animal were examined histomorphometrically. There was an increase in the subepiphyseal cancellous bone trabecullae of the femur. In the growth plate there was an increase in the number of column cells and proliferative cells. These changes were more pronounced in the femur than the humerus. We conclude that swimming induces an increase in subepiphyseal cancellous bone in young adult rats by enhancing growth plate activity.”
Humerus length increased but femoral length decreased.
Articular cartilage height increased in both groups which is very promising:
“During swimming, rats use their hind legs in extensive, forceful upward and downward movements, while the forelegs move in a small circular, balancing fashion. Therefore the effort of the hind legs is greater than that of the forelegs, explaining the differences in the muscle stresses of the humerus and femur and the consequent histomorphological changes in the bones.”
“Growing bones are most responsive to mechanical loading. We investigated bone mass acquisition patterns following a swimming or running exercise intervention of equal duration, in growing rats. We compared changes in bone mineral properties in female Sprague Dawley rats that were divided into three groups: sedentary controls (n = 10), runners (n = 8) and swimmers (n = 11). Runners and swimmers underwent a six week intervention, exercising five days per week, 30min per day. Running rats ran on an inclined treadmill at 0.33 m.s−1, while swimming rats swam in 250C water. Dual energy X-ray absorptiometry scans measuring bone mineral content (BMC), bone mineral density (BMD) and bone area at the femur, lumbar spine and whole body were recorded for all rats before and after the six week intervention. Bone and serum calcium and plasma parathyroid hormone (PTH) concentrations were measured at the end of the 6 weeks. Swimming rats had greater BMC and bone area changes at the femur and lumbar spine (p < 0.05) than the running rats and a greater whole body BMC and bone area to that of control rats (p < 0.05). There were no differences in bone gain between running and sedentary control rats. There was no significant difference in serum or bone calcium or PTH concentrations between the groups of rats. A swimming intervention is able to produce greater beneficial effects on the rat skeleton than no exercise at all, suggesting that the strains associated with swimming may engender a unique mechanical load on the bone.”
“There was no significant difference for femur length between the runners (34.55 ± 1.40mm), swimmers (33.50 ± 0.70mm) or sedentary control rats (34.21 ± 1.99mm, F=1.34, p = 0.28).”<-another study that found shorter femur length for swimming rats. It could be biomechanically upright swimming results in compressive deformation.
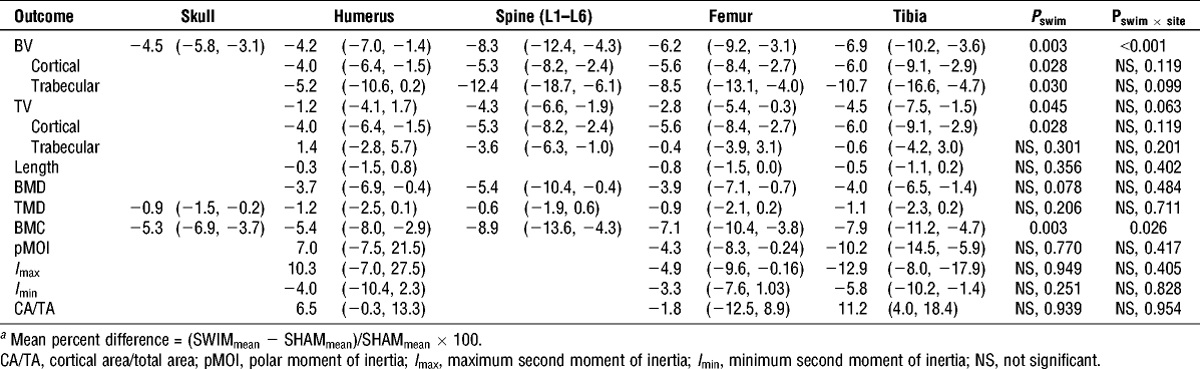
Reduced Bone Mass Accrual in Swim-Trained Prepubertal Mice
“Prepubertal female mice underwent a 16-wk training program, in which they swam for progressively increasing durations up to 55 min for 5 d·wk−1. A sham group was subjected to the water, but they did not perform the swimming exercise. Skeletal sites that were assessed included the proximal humerus, lumbar spine, midshaft and distal femur, proximal tibia, and the skull.”
This study found a decrease in length.
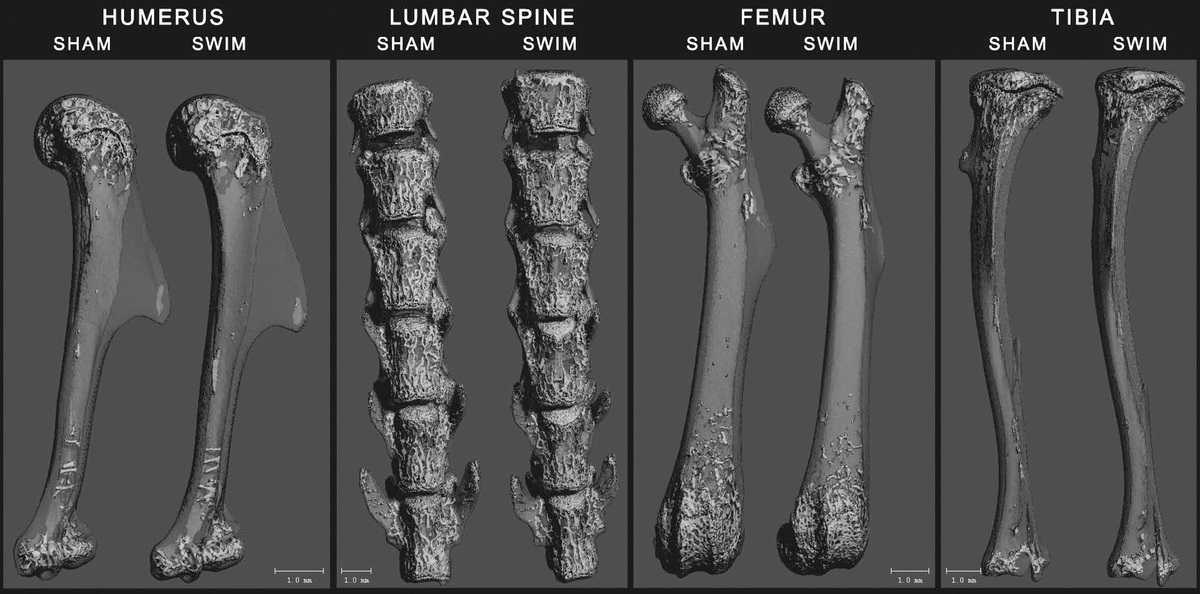
Here are the bone images:
The swimming studies that resulted in growth seem to be swimming studies with attached weight. That may be the key/weight equals more torsion.
It’s possible that a supplement for Neuropeptide Y may not be needed as cold exposure increases Neuropeptide Y levels. But it’s not clear if an increase in Neuropeptide Y will increase height.
Neuropeptide Y Promotes mTORC1 to Regulate Chondrocyte Proliferation and Hypertrophy
“Peripheral neuropeptide Y (NPY) has been reported to regulate bone metabolism and homeostasis, however its potential roles in growth plate chondrogenesis remain unclear. Here, we found that NPY expression decreased during chondrocyte differentiation in vitro and in vivo. NPY was required for chondrocyte proliferation, while in contrast knockdown of NPY facilitated chondrocyte hypertrophic differentiation. {So maybe Neuropeptide Y can keep growth plates open for longer?} Administration of recombinant NPY in rat chondrocytes and metatarsal bones uncoupled the normal proliferation and hypertrophic differentiation during chondrogenesis and thereby inhibited growth plate chondrogenesis and longitudinal bone growth. Remarkably, NPY activated mTORC1 pathway in chondrocytes, whereas attenuation of mTORC1 activity by administration of rapamycin in vitro partially abrogated NPY-mediated effects on chondrocyte proliferation and hypertrophic differentiation. In addition, a combination of Y2R antagonist but not Y1R antagonist with NPY abolished NPY-mediated inhibition of metatarsal growth and growth plate chondrogenesis. Mechanistically, NPY activated Erk1/2 by NPY2R, then phosphorylated ERK1/2 activated mTORC1 to initiate PTHrP expression, which in turn promoted chondrocyte proliferation and inhibited chondrocyte hypertrophic differentiation. In conclusion, our data identified NPY as a crucial regulator of chondrogenesis and may provide a promising therapeutic strategy for skeletal diseases.”
You can potentially buy Neuropeptide Y for lab use. Couldn’t get the full study but here’s another paper on it.
Neuropeptide Y Acts Directly on Cartilage Homeostasis and Exacerbates Progression of Osteoarthritis Through NPY2R
“Neuropeptide Y (NPY) is known to regulate bone homeostasis; however, its functional role as a risk factor during osteoarthritis (OA) remains elusive. In this study, we aim to investigate the direct effect of NPY on degradation of cartilage and progression of OA and explore the molecular events involved. NPY was overexpressed in human OA cartilage accompanied with increased expression of NPY1 receptor (NPY1R) and NPY2 receptor (NPY2R). Stressors such as cold exposure resulted in the peripheral release of NPY from sympathetic nerves{This may explain why people in colder environments are taller}, which in turn promoted upregulation of NPY and NPY2R in articular cartilage in vivo. Intra-articular administration of NPY significantly promoted chondrocyte hypertrophy and cartilage matrix degradation, with a higher OARSI score than that of control mice, whereas inhibition of NPY2R but not NPY1R with its specific antagonist remarkably ameliorated NPY-mediated effects. Moreover, NPY activated mTORC1 pathway in articular chondrocytes, whereas the administration of rapamycin (an mTORC1 inhibitor) in vitro abrogated NPY-mediated effects. Mechanistically, mTORC1 downstream kinase S6K1 interacted with and phosphorylated SMAD1/5/8 and promoted SMAD4 nuclear translocation, resulting in upregulation of Runx2 expression to promote chondrocyte hypertrophy and cartilage degradation. In conclusion, our findings provided the direct evidence and the crucial role of NPY in cartilage homeostasis.”
“peripheral NPY can be also synthesized by osteoblasts, osteocytes, as well as chondrocytes both at embryonic and adult stage”
“the NPY-treated group exhibited more severe OA phenotypes than that of the control group as indicated by significant cartilage degradation and fibrillation together with a higher OARSI score”<-although we don’t know how that will translate to height.
“NPY activated mTORC1 signaling in articular cartilage, which in turn activated SMAD1/5/8 signaling and consequently promoted chondrocyte hypertrophy and cartilage degradation.”
“old exposure stimulates NPY release from sympathetic nerves, and then NPY could enter the blood since NPY-positive sympathetic nerve fibers have been found to be mostly distributed around and within blood vessels. mice exposed in cold water increased circulating NPY.”
Skeletal phenotype of the neuropeptide Y knockout mouse
“lobal deletion of NPY results in a smaller femoral cortical cross-sectional area (-12%) and reduced bone strength (-18%) in male mice. In vitro, NPY-deficient bone marrow stromal cells (BMSCs) showed increase in osteogenic differentiation detected by increases in alkaline phosphatase staining and bone sialoprotein and osteocalcin expression. Despite both sexes presenting with increased adiposity, female mice had no alterations in bone mass, suggesting that NPY may have sex-specific effects on bone”
Neuropeptide Y Regulation of Energy Partitioning and Bone Mass During Cold Exposure
“wild-type (WT) mice at thermoneutral (29 °C) were compared to mice at room temperature (22 °C) conditions. Interestingly, bone mass was lower in cold-stressed WT mice with significant reductions in femoral bone mineral content (− 19%) and bone volume (− 13%).”
“It is a common observation that normal growth is affected or suppressed in young individuals by environmental adversities and physical disturbances. On the other hand, when health or favorable conditions are restored, the tempo of growth promptly accelerates as if the individual is trying to make up for the growth debt incurred.”
“Growth retardation during the single cold exposure period, and prompt compensatory growth after return to the warm environment, occurred in both the tail and the body, and in the younger A as well as the older B rats. During the 2-week periods of cold exposure, tail gain of the A rats was only about 30 % of that of the warm controls (W2), while that of the B rats averaged about 50 %. Likewise, the rapid compensatory growth in tail length during the 2 weeks immediately after the end of cold exposure was more prominent in the A than in the B rats: the tail gain in the A rats averaged around 145 % of that of the controls in contrast to 130 % in the B rats. Not only was the compensation greater in the A rats, but the compensatory period (period of significantly faster growth) was longer also, extending over 8 weeks (from 3rd to 10th), in contrast to 4 weeks (from 6th to 10th) in the B rats. By the end of the 10th week the tail length of A and B rats was not significantly shorter than that of the W2 group”
Uncoupling protein-1 is protective of bone mass under mild cold stress conditions
“UCP-1 is critical for protecting bone mass in mice under conditions of permanent mild cold stress for this species (22 °C). UCP-1−/− mice housed at 22 °C showed significantly lower cancellous bone mass, with lower trabecular number and thickness, a lower bone formation rate and mineralising surface, but unaltered osteoclast number, compared to wild type mice housed at the same temperature. UCP-1−/− mice also displayed shorter femurs than wild types, with smaller cortical periosteal and endocortical perimeters. Importantly, these altered bone phenotypes were not observed when UCP-1−/− and wild type mice were housed in thermo-neutral conditions (29 °C), indicating a UCP-1 dependent support of bone mass and bone formation at the lower temperature. Furthermore, at 22 °C UCP-1−/− mice showed elevated hypothalamic expression of neuropeptide Y (NPY) relative to wild type, which is consistent with the lower bone formation and mass of UCP-1−/− mice at 22 °C caused by the catabolic effects of hypothalamic NPY-induced SNS modulation.”