It’s long been noticed that heat can increase longitudinal bone growth but mostly there has been a lack of concrete numbers. It may have little impact on adult height growth due to mostly impacting the growth plate however heat may have an impact on stem cells.
Unilateral Heat Accelerates Bone Elongation and Lengthens Extremities of Growing Mice
“Current bone lengthening protocols involve invasive surgeries or drug regimens, which are only partially effective. Exposure to warm ambient temperature during growth increases limb length, suggesting that targeted heat could noninvasively enhance bone elongation. Daily heat exposure on one side of the body unilaterally increases femoral and tibial lengths. Mice (N = 20) were treated with 40C unilateral[only one side of the body] heat for 40 minutes/day for 14 days post-weaning[after no longer taking breast milk]. Non-treated mice(N = 6) served as controls. Unilateral increases in ear (8.8%), hindfoot (3.5%), femoral (1.3%) and tibial(1.5%) lengths were obtained.Tibial elongation rate was >12% greater (15 µm/day) on the heat-treated side.Extremity lengthening correlated with temperature during treatment. Body mass and humeral length{It’s interesting that heat only worked for certain bones} were unaffected. To test whether differences persisted in adults, mice were examined 7-weeks post-treatment. Ear area, hindfoot, femoral, and tibial lengths were still significantly increased ∼6%, 3.5%, 1%, and 1%, respectively, on the heat-treated side{So the heat treatment permanently increased growth and not just growth rate. That ear area increased shows that the cartilage itself is affected}. Left-right differences were absent in non-treated controls, ruling out inherent side asymmetry. This model is important for designing noninvasive heat-based therapies to potentially combat a range of debilitating growth impediments in children.”
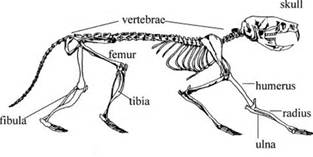
Here’s a rodent skeleton:
Let’s say your tibia and femur together are about 30 inches. A one percent increase would be about .3inches resulting in a 3rd of an inch increase in height. Since the heat applied is not optimized there’s the possibility for more.
“A major obstacle to successful bone lengthening by noninvasive means is difficulty in targeting therapeutics to cartilaginous growth plates, which do not have a direct blood supply{Note that LSJL is able to target the growth plates and the stem cells within the growth plates via mechanical loading}. Experimental drug delivery approaches include surgically implanted catheters and localized injections into specific growth plates”<-Note that Ping Zhang was able to stimulate length growth without injecting directly into the growth plate via localized IGF2 injections.
“Data from our lab and others demonstrate that exposure to warm ambient temperature
during growth increases bone blood supply and length in young mice.”<-The increase in bone blood supply could have an impact in adults as well if the right stimuli was in place to induce the formation of new growth plates.
“intermittent targeted heating could be accomplished with a heating pad or temperature cuff.”
Here’s an adjustable heating pad. I haven’t testing it myself and I’m not sure if it’s the best one but it is a sample:
“goal is to develop a low-cost, noninvasive method for lengthening bones that can translate
into practical therapy to offset linear growth impediments in children.”<-They believe that heat can work in human children to increase growth.
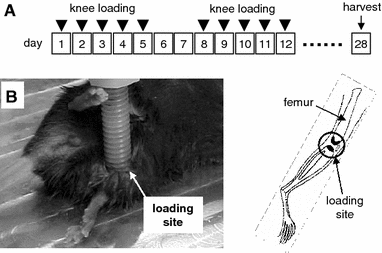
“Mice were anesthetized with 1.5% isoflurane and placed in lateral recumbency on a
40C heating pad for 40 minutes each day”
“The 3-5 week age interval is a time of rapid, temperature-sensitive growth in mice. By comparison, this period could be considered roughly similar to human development between toddler age and entry to middle school.”
“Limbs on the heat-treated side were wrapped in custom fitting thermal booties to ensure uniform heat distribution”
“In addition to limb length, cartilaginous ears were measured to document a treatment effect because ear size increases with ambient temperature”
“Skin temperatures of heat-treated hindfeet and ears averaged 40C during treatments. Nontreated side temperatures averaged 30C with no major fluctuations”<-So 10C of heat increase resulted in a 1% increase in leg length. It’s unclear whether more heat would result in more lengthening.
“When nontreated and heat-treated sides were analyzed in aggregate, there were significant positive correlations between hindlimb temperature and tibial elongation rate”
“short-term (30-minute) hindlimb heating increases molecular uptake in mouse tibial growth plates”<-this doesn’t really help adults create neo-growth plates.
“Although it is unclear why humeral length did not differ, one potential explanation is the warmer starting temperature of the forelimb when compared to the hindlimb. The knee joint capsule is normally at least 3-4C lower than body core.Our treatments elevated hindlimb temperature by 10C on the heat-treated side.
However, skin temperatures in the humeral region more closely resembled body core, consistent with thermal maps for humans showing that 37C core temperature extends into the shoulder region, while extremity temperatures progressively decrease in a proximal-distal gradient.”
“Our working hypothesis is that heat-treatments do not impact temperature of the humerus
due to the proximity of this joint to the body core, and the disproportionately large volume of
warm blood delivered to the shoulder region through the large subscapular artery”
“the left-right symmetry in humeral length reflects its relatively constant temperature. This could be tested by decreasing shoulder temperature with cold, which stunts limb elongation in a dose-dependent manner”
“up to a 20% increase in bone growth rate for every one degree C increase in incubation temperature in growing chicks.”
“unilateral exposure of mild, non-damaging 40C heat for 40-minutes per day for only 14 days permanently increased ear area and hindlimb length on heat-treated sides of young mice.”
“the width of the mouse growth plate is only a fraction of that of a human growth plate. It will be important to replicate these results in a larger animal model to ensure that heat can fully penetrate a larger growth plate, so as to avoid potential angular growth deformities. This should not be problematic, however, sincewhole body heat-effects on bone length have already been demonstrated in experiments using large animals”
I already feel a bit of a temperature elevation when performing LSJL.
According to Periodic heat shock accelerated the chondrogenic differentiation of human mesenchymal stem cells in pellet culture., Heat increased chondrogenic differentiation although they were cells already committed to the chondrogenic lineage. 41C for 1 hour was the stimulus used.
Hindlimb heating increases vascular access of large molecules to murine tibial growth plates measured by in vivo multiphoton imaging.
“Advances in understanding the molecular regulation of longitudinal growth have led to development of novel drug therapies for growth plate disorders. Despite progress, a major unmet challenge is delivering therapeutic agents to avascular-cartilage plates. Dense extracellular matrix and lack of penetrating blood vessels create a semipermeable “barrier,” which hinders molecular transport at the vascular-cartilage interface. To overcome this obstacle, we used a hindlimb heating model to manipulate bone circulation in 5-wk-old female mice (n = 22). Temperatures represented a physiological range of normal human knee joints. We used in vivo multiphoton microscopy to quantify temperature-enhanced delivery of large molecules into tibial growth plates. We tested the hypothesis that increasing hindlimb temperature from 22°C to 34°C increases vascular access of large systemic molecules{This is a limited temperature range so we don’t know if there’s a cap on how much temperature can increase height}, modeled using 10, 40, and 70 kDa dextrans that approximate sizes of physiological regulators. Vascular access was quantified by vessel diameter, velocity, and dextran leakage from subperichondrial plexus vessels and accumulation in growth plate cartilage. Growth plate entry of 10 kDa dextrans increased >150% at 34°C. Entry of 40 and 70 kDa dextrans increased <50%, suggesting a size-dependent temperature enhancement. Total dextran levels in the plexus increased at 34°C, but relative leakage out of vessels was not temperature dependent. Blood velocity and vessel diameter increased 118% and 31%, respectively, at 34°C. These results demonstrate that heat enhances vascular carrying capacity and bioavailability of large molecules around growth plates, suggesting that temperature could be a noninvasive strategy for modulating delivery of therapeutics to impaired growth plates of children.”
“transporting agents into and through the dense ECM of cartilage remains a challenge.”
“There are two components of the vascular-cartilage barrier that determine solute availability in growth plates: 1) ability of molecules to exit the vasculature and 2) ability to enter the cartilage matrix. The endpoint of interest is total uptake in the growth plate, since this is what ultimately impacts chondrocyte performance.”
“heat increases small solute (<500 Da) uptake in growth plate cartilage in vivo. In the current study, we sought to determine whether heat could increase uptake of large molecules (≥10 kDa).”
” Healthy human knee joint temperatures can range from under 20°C to over 37°C during normal and clinical activities. By modulating limb temperature within this natural range, we show that mild 34°C heating increases transport of large (≥10 kDa) molecules that are otherwise size restricted at the vascular-cartilage interface.”<-So this study doesn’t tell us whether non-physiological temperatures can help.
“The three principal vascular routes by which blood reaches the growth plate, are epiphyseal vessels, metaphyseal vessels, and a circumferential ring vessel connected to a subperichondrial plexus. The epiphyseal and metaphyseal vessels course within the bone, whereas the circumferential vessel and its plexus are located just deep to the perichondrium (i.e., subperichondrial) that surrounds the cartilage plate. Soluble factors reach the growth plate through all three vascular routes, and temperature can affect blood flow through these vessels”
“Heat increases vessel diameter and blood velocity in subperichondrial plexus vessels.”

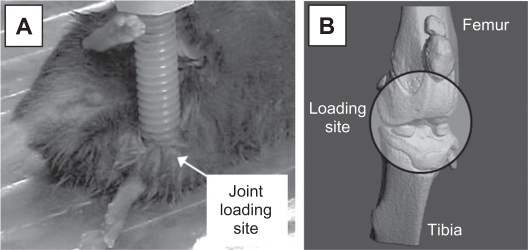
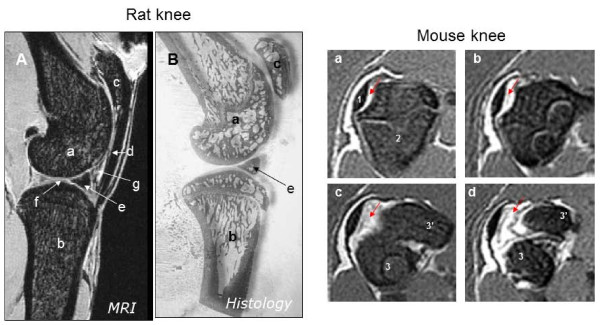


 Hypothesized theory of input of compression and tension on growth rate.
Hypothesized theory of input of compression and tension on growth rate.